- Review
- Open access
- Published:
Hydroxylases involved in terpenoid biosynthesis: a review
Bioresources and Bioprocessing volume 10, Article number: 39 (2023)
Abstract
Terpenoids are pervasive in nature and display an immense structural diversity. As the largest category of plant secondary metabolites, terpenoids have important socioeconomic value in the fields of pharmaceuticals, spices, and food manufacturing. The biosynthesis of terpenoid skeletons has made great progress, but the subsequent modifications of the terpenoid framework are poorly understood, especially for the functionalization of inert carbon skeleton usually catalyzed by hydroxylases. Hydroxylase is a class of enzymes that plays an important role in the modification of terpenoid backbone. This review article outlines the research progress in the identification, molecular modification, and functional expression of this class of enzymes in the past decade, which are profitable for the discovery, engineering, and application of more hydroxylases involved in the plant secondary metabolism.
Graphical Abstract

Introduction
Terpenoids are the most diverse and abundant class of secondary metabolites among natural products, featuring tremendous structural and functional diversity with a series of significant pharmacological and biological activities (Chacon-Morales 2022). More than 80,000 terpenoids have been recorded in the Dictionary of Natural Products (http://dnp.chemnetbase.com) (Chen et al. 2020).
The extraordinary low content of these terpenoids in natural resources renders a direct extraction unsustainable. Moreover, chemical syntheses of complex terpenoids are generally received with the tedious multiple steps and extremely low overall yield. In addition, a large number of organic reagents are required for the above two approaches to ensure the extraction rate and product purity, posing serious circumstance destructions. In the past decade, the rapid developments in synthetic biology offer new possibilities for the efficient heterogeneous overproduction of terpenoids in microbial hosts, which can reduce the use of organic reagents and shorten the synthetic routes to a certain extent. At present, many kinds of terpenoids have been synthesized by synthetic biological methods, such as antimalarial compound artemisinin (Paddon and Keasling 2014), the precursor of famous anticancer drugs taxol (Ajikumar et al. 2010; Walls et al. 2021), and liver protecting ingredient glycyrrhizic acid (Sun et al. 2020).
Heterogeneous biosynthesis of terpenoids depends on the clarification of the corresponding metabolic pathways. In the biosynthetic pathway of terpenoids, two kinds of C5 building blocks, isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAPP), are cyclized or arranged to form different types of oligomers, such as geranyl pyrophosphate (GPP), farnesyl pyrophosphate (FPP), and geranylgeranyl pyrophosphate (GGPP). Then, a series of terpenoid skeletons are assembled under the catalysis of various cyclases. The subsequent modification processes of these skeletons contribute greatly to the structural diversity and pharmacological activities of terpenoids, which are mainly enabled by cytochrome P450 enzymes (CYPs) (Behrendorff and Gillam 2017).
CYPs are able to activate the inert C–H bond on the terpene backbone with high stereoselectivity, which keep been the bottlenecks to breakthrough in chemical synthesis. Although CYPs are capable to catalyze a variety of reactions, including hydroxylation, dealkylation, sulfur oxidation, epoxidation, deamination, desulfurization and peroxidation reactions, the most iconic reaction remains the insertion of hydroxyl or oxygen atoms into the C–H backbone to form the corresponding alcohols, which may be further oxidized to aldehydes or acids. These oxygen-containing groups are often necessary for the pharmacological activity of natural products. Harvesting the capacity of hydroxylases would be anticipated to greatly enhance the producibility of valuable natural products. However, some CYPs exhibit the low heterologous expression, poor stability, low catalytic activity, as well as the catalytic promiscuity of the extant CYPs, representing the bottlenecks of terpenoids heterologous biosynthesis.
This article reviews the various types of hydroxylases, especially cytochrome P450s, involved in the skeleton modifications of monoterpenes, sesquiterpenes, diterpenes, triterpenes, and tetraterpenes, and their corresponding oxidations. The challenges in the current research and corresponding measures are summarized. We expect that this summary can provide a guidance for the discovery of new terpenoid hydroxylases and the subsequent heterologous expression and modifications to exploit novel bioactive compounds for medicinal applications.
Classification of terpenoid hydroxylases
The principle of terpenoid biosynthesis is using terpene synthase to unite the isoprene subunit chain to form monoterpenes, sesquiterpenes, diterpenes, triterpenes, and tetraterpenes according to the number of isoprene units (Huang et al. 2021). Terpenoid hydroxylases then catalyze the oxidation to activate the different carbon skeletons. Based on the different substrates, hydroxylases are classified into the following four categories.
Monoterpene hydroxylases
Monoterpenes refer to terpenes and their derivatives containing two isoprene units in molecules, which are widely found in the secreted tissues of higher plants. Most of them have strong aroma and physiological activity and are important raw materials for the pharmaceutical and cosmetic industries. Monoterpene oxidases had been found in plants, microorganisms, and animals. According to the diversity of skeleton structure, monoterpene skeletons can be divided into acyclic and monocyclic forms (Corinna and Henrik 2013).
The CYP76 family occupies a large proportion in the monoterpene hydroxylases which catalyze the allylic hydroxylation of acyclic monoterpenes. For example, CYP76C1, CYP76C2, CYP76C4, and CYP76B6 can realize the allylic hydroxylation of linalool, geraniol, nerol, and citronellol (Hofer et al. 2014). CYP76C2 and CYP76C4 can also catalyze epoxidation of linalool at C-1 and C-2. CYP71B31 and CYP76C3, identified in Arabidopsis thaliana (A. thaliana), can also catalyze hydroxylation of linalool at multiple sites and epoxidation at C-1 and C-2 to produce four kinds of products, respectively (Ginglinger et al. 2013). Geraniol is the main agent of rose flavor essence and its derivatization may create novel flavor components. CYP76F45 can simultaneously catalyze allylic hydroxylation of monoterpene geraniol, sesquiterpene farnesol, and diterpene geranylgeraniol (Sintupachee et al. 2015). In recent years, P450s identified in beetles have attracted the attention of researchers. The pine bark beetle can convert myrcene into ipsdienol, the aggregation pheromone component. There are at least seven P450s in the organs or tissues of the mountain pine beetle. As the first cytochrome P450 known from insect, CYP6BH5 was identified from the liposome of leaf beetle and was expressed in Sf9 insect cells, and then catalyzed the hydroxylation of geraniol and its structural analogs, nerol and citronellol (Fu et al. 2019).
Limonene is a widely studied monocyclic monoterpene. Many alkane degrading microorganisms can catalyze L-limonene to produce perillyl alcohol. A limonene 7-hydroxylase CYP153A6 was isolated from the Mycobacterium SP.HXN-1500 strain (van Beilen et al. 2005). CYP750B1 and CYP76AA25 identified from Thuja Plicata can catalyze the allylic hydroxylation of dicyclic monoterpene (+)-sabinene to produce (+)-trans-sabinol which is the biosynthetic precursor of α-thujone. The substrate spectrum of CYP76AA25 is wide and also applicable to sesquiterpenoid farnesene and herbicide isoproturon (Gesell et al. 2015). CYP345E2 (Keeling et al. 2013), CYP6DE1 (Chiu et al. 2019a), CYP6DE3 (Nadeau et al. 2017), and CYP6DJ1 (Chiu et al. 2019b) were identified from mountain pine beetle and had been functional characterized to enable the allylic hydroxylation of α-pinene, β-pinene, limonene, and other monoterpenes to produce a variety of products with significant application value (Song et al. 2013). The difference is that CYP6DJ1 is specified to the two enantiomers of monocyclic monoterpene limonene to generate various epoxides and perillyl alcohol, while it has no catalytic activity for dicyclic monoterpene pinene and other terpinolenes.
Vindoline is a kind of tabersonine-derived monoterpene indole alkaloid (MIA) from Catharanthus roseus (C. roseus), and its main role is to resist the ingestion of herbivores to protect plants. Vindoline serves as the precursor to vinblastine and vincristine which are used as antitumor drugs. CYP71D12 and CYP71D351 were found to hydroxylate the C-16 site of tabersonine with extremely high affinity and substrate specificity (Besseau et al. 2013). CYP71BJ1 promotes the hydroxylation of the C19 site in tabersonine to produce 19-hydroxytabersonine, which is then further epoxidized by 6,7-epoxidase to generate 19-O-acetylhoerhammericine (Giddings et al. 2011) (Fig. 1). Due to that CYP71BJ1 can also adopt lochnericine and tabersonine as substrates in vitro, the detailed pathway of hoerhammericine biosynthesis remains unclear (Fig. 2).

Proposed biosynthetic pathway of tabersonine-derived alkaloids (vindoline and 19-O-acetylhoerhammericine) in C. roseus

Monoterpene hydroxylases involved in this article
Sesquiterpene hydroxylases
Sesquiterpenoids are natural terpenoids with higher boiling points than monoterpenes, which are polymerized from three molecules of isoprene. Most of their oxygen-containing derivatives have strong aroma and obvious biological activity. Sesquiterpene displays highly diverse branched skeletons, including mono-(Germacrene A, humulene, and zerumbone), bi-(β-caryophyllene, amorpha-4,11-diene, (+)-valencene, (+)-nootkatone), and tricyclic hydrocarbon skeletons (pentalenene, α-cedrene, and (+)-3(15)-cedren-4-ol) (Corinna and Henrik 2013).
Sesquiterpene lactones (STLs) are unique natural metabolites in Asteraceae plants. They can be used as anti-inflammatory, sedative, analgesic, anticancer, and anti-malaria drugs. Germacrene A (GA) and germacra-1(10),4,11(13)-trien-12-oic acid (GAA) are important monocyclic precursors to the synthesis of sesquiterpene lactones. CYP71AV2 has the function as GA oxidase, which promotes the oxidation reaction of the methyl group at C-12 of GA to generate GAA in three consecutive steps (Ramirez et al. 2013). Ikezawa et al. identified GAA-6-hydroxylase CYP71BL2 and GAA-8-hydroxylase CYP71BL1 from lettuce and sunflower, respectively (Ikezawa et al. 2011). Eljounaidi et al. isolated GA hydroxylase CYP71AV9 and GAA hydroxylase CYP71BL5 from Cynara cardunculus (Eljounaidi et al. 2014). Co-expression of CYP71AV9 together with GA synthase (GAS) during the biosynthesis of germacra-1(10),4,11(13)-trien-12-oic acid in yeast indicated that CYP71AV9 enabled the hydroxylation of position C-12 of GA, with alcohols and aldehydes generated under acidic condition and acids under buffer culture condition (Eljounaidi et al. 2014). The co-expression of CYP71BL5 and CYP71AV9 with GAS in Nicotiana benthamiana (N. benthamiana) leads to costunolide conjugates verified CYP71BL5 as the costunolide synthase (Fig. 3).

Proposed biosynthetic routes to C6–C7 and C7–C8 sesquiterpene lactones
Monocyclic sesquiterpene zerumbone compounds exhibit anticancer, anti-inflammatory, and antinociceptive activities for potential application (Chien et al. 2016). The biosynthesis of zerumbone includes the selective hydroxylation of the basic skeleton α-humulene by cytochrome P450 to produce 8-hydroxy-humulene and then releases zerumbone through the alcohol dehydrogenase. Schifrin et al. isolated two sesquiterpene hydroxylases CYP260A1 and CYP264B1 from Sorangium cellulosum (S. cellulosum) Soce56 (Schifrin et al. 2015). CYP260A1 was found to catalyze the hydroxylation of monocyclic sesquiterpenes zerumbone (b) and bicyclic sesquiterpenes (+)-nootkatone (d), (+)-3(15)-cedren-4-ol (g) to form a variety of oxidation products. Compared to CYP260A1, CYP264B1 has a wider substrate tolerance, including monocyclic compounds zerumbone and α-humulene (a) and more complicated compounds, such as (+)-nootkatone, (+)-valencene (c), β-caryophyllene (e), and α-cedrene (f) (Fig. 4). It is worth noting that compared with CYP260A1, CYP264B1 has higher regioselectivity, with fewer kinds of products generated and a high proportion of specific product. The substrate promiscuity of these two hydroxylases is continually expanded and improved through molecular modification in order to produce the required high value-added products.

Substrate spectrum of CYP260A1 and CYP264B1
Bicyclic sesquiterpene artemisinin and derivatives represent the most effective class of antimalarial drugs. Its biosynthetic pathway has been largely elucidated in the past two decades. Firstly, farnesyl diphosphate is cyclized to amorpha-4,11-diene by amorphadiene synthase, and then CYP71AV1 catalyzes the three-step continuous hydroxylation of amorphadiene (Fig. 5). The regioselective oxidation at position C-12 produces artemisinol and that is further converted to artemisinic acid (Wang et al. 2013). The sequence identity of GA oxidase CYP71AV2 and amorpha-4,11-diene monooxygenase CYP71AV1 is as high as 86.7%, indicating that these two oxygenases may be evolved from the same ancestor.

Biosynthetic pathway for sesquiterpene lactones artemisinin and arteannuin B
Nootkatone, a bicyclic sesquiterpene compound with a grapefruit-like odor, is widely used in the food and fragrance industries. The high cost of natural extraction limits its industrial application. Cankar et al. screened out a new P450 protein CYP71AV8 with the sequence identity of 81% to amorpha-4,11-diene oxidase CYP71AV1. CYP71AV8 was able to catalyze the hydroxylation of (+)-valencene at the C-2 position to yield (+)-nootkatol, and a small part was further oxidized to (+)-nootkatone. With additional activities, like GA oxidase and amorphadiene oxidase, CYP71AV8 promotes the sequential oxidation of both at the C-12 position to the corresponding alcohols, aldehydes, and acids. This is the first P450 that has been proven to have the hydroxylation function of (+)-valencene (Cankar et al. 2011). Girhard et al. verified that CYP109B1 from Bacillus subtilis can also catalyze the sequential oxidation of valencene at the C-2 position to produce nootkatone (Girhard et al. 2010). Takase et al. used homologous sequencing to identify CYP71BE5, which catalyzes the hydroxylation of α-guaiene at C-2 to generate alcohol, and further oxidation produces (−)-rotundone. This enzyme can also catalyze the hydroxylation of (+)-valencene at C-2 to yield β-nootkatol (Takase et al. 2016). Gavira et al. also isolated CYP71D51V2 from Saccharomyces cerevisiae (S. cerevisiae) to enable the oxidation of (+)-valencene to (+)-nootkatol as a major product (Gavira et al. 2013) (Fig. 6).

Conversion of (+)-valencene to (+)-nootkatone via trans- and cis-nootkatol catalyzed by CYP71AV8
Members of the genus Streptomyces produce a variety of biologically active secondary metabolites, including most of the antibiotics widely used in medicine. A few of terpene hydroxylases were identified in the gene cluster of antibiotics biosynthesis. Quaderer et al. found CYP183A1 in the gene cluster for the biosynthesis of the tricyclic sesquiterpene antibiotic pentalenolactone catalyzes the hydroxylation of pentalenene at the allylic position to generate pentalen-13-ol, which is then gradually oxidized to pentalen-13-al, the precursor toward antibiotic pentalenolactone (Quaderer et al. 2006). Zhao et al. identified gene of CYP170A1 in Streptomyces coelicolor A3(2) and confirmed that CYP170A1 catalyzes the two-step continuous allylic oxidation of epi-isozizaene to generate the sesquiterpene antibiotic albaflavenone (Zhao et al. 2008) (Fig. 7). CYP170A1 was also found to convert farnesyl diphosphate (FDP) into acyclic sesquiterpene farnesene and farnesol. Enzymology and structural data clearly showed that CYP170A1 has active sites of farnesene synthase, which make it perform the additional terpene synthase activity in the presence of divalent cations (Zhao et al. 2009) (Fig. 8).

Bifunctional CYP170A1 with two different active sites. a Allylic oxidation of epi-isozizaene; b farnesyl diphosphate into acyclic sesquiterpene farnesene, farnesol, and nerolidol

Sesquiterpene hydroxylases involved in this article
Diterpene hydroxylases
Diterpenes are higher terpenoids polymerized from four molecules of isoprene. Due to their large molecular weight and poor volatility, diterpenes mostly exist in nature in the form of resins and lactones. There are very few acyclic and monocyclic diterpenoids in plants, mainly bicyclic and tricyclic diterpenoids, especially numerous oxygen-containing derivatives.
Cembranoids contain a large group of 14-membered macrocyclic diterpenoids whose types, positions, and stereochemical properties of their oxygen function are different, which are mainly derived from tobacco and corals. Cembranoids have a variety of biological activities, including antitumor and neuroprotective effects (Schrepfer et al. 2020). There exist narrow reports on CYP450s involved in their biosynthesis pathway. Wang et al. isolated the gene of CYP71D16 from trichome glands of tobacco using PCR-based complementary cDNA subtraction strategy. CYP71D16 catalyzed the hydroxylation at C-6 of cembratriene-ol (CBT-ol) to produce cembratriene-diol (CBT-diol), which is a key diterpene in the biosynthesis of the other cembranic compounds (Wang and Wagner 2003) (Fig. 9).

Biosynthetic pathways of CBT-ol and CBT-diol in tobacco. GGPP geranylgeranyl pyrophosphate
By constructing P450 BM3 mutant library, Le-Huu et al. obtained a double P450 BM3 mutant F87A/I263L oxidize β-CBT-diol at C-9 with 100% regioselectivity and 89:11 diastereomeric ratio. A triple mutant L75A/V78A/F87G oxidize β-CBT-diol at C-10 with 97% regioselectivity and the diastereomeric ratio was 74:26 (Le-Huu et al. 2015). Following research showed that the triple mutant L75A/V78A/F87G can also hydroxylase C-10 of (9R)-β-CBT-triol to form (9S,10S)-β-CBT-traol and (9S,10R)-β-CBT-traol with diastereomeric ratio of 90:10 (Le-Huu et al. 2016) (Fig. 10).

a Hydroxylation of β-CBT-diol catalyzed by P450 BM3 variants; b one-pot two-step hydroxylation of β-CBT-diol to form β-CBT-traol catalyzed by P450 BM3 variants. 8: (9R/S)-β-CBT-triols; 9: (10R/S)-β-CBT-traols; 10: (9S,10R/S)-β-CBT-traols
The products derived from the casbene and the related derivatives are part of the most representative bicyclic diterpenes, exhibiting significant antitumor and antiviral activities. King et al. obtained casbene synthases and various P450s belonging to CYP726A through gene cluster mining. CYP726A14, CYP726A17, or CYP726A18 from Ricinus communis (R. communis) can catalyze the 5-oxidation of casbene to 5-keto-casbene and 5α-hydroxy-casbene. CYP726A16 was not able to functionalize casbene. However, by co-expressing casbene synthase, casbene-5-oxidase, and CYP726A16, 5-keto-7,8-epoxy-casbene was able to detected, indicating that the conversion of casbene to 5-keto-casbene is a linear step in the diterpenoid biosynthetic pathway. Interestingly, CYP726A15 derived from the same gene cluster can catalyze the oxidation of the C-5 position of neocembrene in two consecutive steps to produce 5-keto-neocembrene. Additionally, CYP726A19 from Euphorbia peplus (E. peplus) has the function of casbene 5-oxidase to give the same products as that used by CYP726A14 with a different proportion of the hydroxy- and the keto-products (King et al. 2014). The mature seeds of Euphorbia lathyris L. contain many macrocyclic diterpenes. Through transcriptome analysis of the mature seeds of Euphorbia lathyris L. and intracellular functional characterization, Luo et al. discovered two P450s function as regiospecific casbene hydroxylases, CYP71D445 and CYP726A27, that can catalyze the continuous two-step oxidation of C-9 and C-5 of casbene, respectively.
Some of these enzymes are bifunctional enzymes. The initial product by CYP726A14 was characterized to be 5-hydroxy-casbene, which was subsequently oxidized to 5-keto-casbene and 5-keto-6-hydroxy-casbene with further hydroxylation (Boutanaev et al 2015). The combined in vitro assays of CYP71D445 and CYP726A27 demonstrated that the two P450s act as bifunctional enzymes, thereby clarifying their shared responsibility with a partial functional redundancy for catalyzing oxidation at C-6. Multifunction of these hydroxylases plays an important role in the diterpene modification leading to a great structural diversification (Fig. 11).

a Reactions of casbene catalyzed by CYP726A14 and potential role in castor bean diterpenoid biosynthesis. b Proposed pathway for the conversion of casbene to casbene-5,6,9-triol through enzymatic hydroxylations catalyzed by CYP71D445 and CYP726A27
One of the representative tricyclic diterpenoids is carnosic acid and its related derivatives derived from the Lamiaceae plants. Those diterpenoids exhibit a variety of effects, including acting as anti-HIV and anticancer agents. The cyclized product miltiradiene from GGPP by the diterpene synthase is then oxidized to abietatriene and finally hydroxylated by a series of effective P450s, such as CYP76AH1 from Salvia miltiorrhiza (S. miltiorrhiza), CYP76AH4–7 from Salvia fruticosa (S. fruticosa) and CYP76AH22–24 from Rosmarinus officinalis (R. officinalis). With the initial name as ferruginol synthases (FSs), these P450s can hydroxylate dehydroabietadiene or abietatriene at C-12 to produce ferruginol and then play the role of the ferruginol 11-hydroxylase (HFSs) to generate 11-hydroxyferruginol (Bozic et al. 2015; Scheler et al. 2016; Zi and Peters 2013). Moreover, CYP76AK6–8 from R. officinalis and S. fruticosa can perform the three-step continuous oxidation on the C-20 position of CYP76AK6–8 to give carnosic acid. In addition, CYP76AK6–8 have the catalytic activity of oxidation on a variety of substrates, such as a direct oxidation of miltiradiene at the C-20 position to yield miltiradiene 20-ol. The discovery of a series of CYP450 genes involved in the biosynthetic pathway of carnosic acid was realized by the heterologous synthesis in yeast, providing an elegant example for the de novo synthesis of carnosic acid in other organisms, such as plants (Scheler et al. 2016).
The promiscuity of some oxidases involved in this pathway leads to a dispersion of metabolic flow. CYP76AH3 can catalyze the oxidation of ferruginol at positions of C-11 and C-7 to deliver two intermediates, 11-hydroxy ferruginol, and 11-hydroxy sugiol, with no strict order of hydroxylation. CYP76AK1 was characterized to continue the transformation of the two intermediates produced by CYP76AH3, and hydroxylation of the C-20 position to form 11,20-dihydroxy ferruginol and 11,20-dihydroxy sugiol as the endpoint. Among them, the C-11 and C-12 hydroxyl groups of 11,20-dihydroxy ferruginol can be spontaneously oxidized to form the carbonyl groups in 10-hydroxymethyl tetrahydromiltirone (Guo et al. 2016). The catalytic promiscuity of CYP76AH3 and CYP76AK1 resulted in a variety of intermediates within the metabolic process (Fig. 12).

Proposed partial biosynthetic pathway of carnosic acid, carnosol and 11,20-dihydroxy sugiol. FSs: ferruginol synthases, including CYP76AH1, CYP76AH3, CYP76AH4–7, and CYP76AH22–24. HFSs: 11-hydroxyferruginol synthases
Resin acid is an important defensive component of conifers and has a protective effect against rapidly evolving pests and pathogens. A variety of diterpene cyclases in conifers catalyze GGPP to generate different products, including pimaradiene, isopimaradiene, sandaracopimaradiene, and dehydroabietadiene. CYP720B1 from Pinus taeda (P. taeda) and CYP720B4 from Picea sitchensis (P. sitchensis) can oxidize the C-18 sites of the above four substances in three consecutive steps to release the corresponding resin acids. The sequence identity of these two P450s is 86%, and the sequence similarity is 95%. Among them, CYP720B4 expressed in Escherichia coli (E. coli) and engineered yeast shows that its hydroxylation activities on multiple substrates produce various oxidized diterpenes (Hamberger et al. 2011).
Taxol (Paclitaxel®) has low toxicity and potent anticancer activity and has been widely studied because of its extremely high application value. Taxadiene cyclase effectively converts GGPP to taxadiene, which is then hydroxylated by a series of P450s. Up to now, the complicated consecutive oxidation process catalyzed by various P450s has not been fully resolved. The first step is generally adapted to be the hydroxylation of the C-5 position of taxadiene by CYP725A4 to form taxadiene-5α-ol. The heterologous expression of CYP725A4 in E. coli and S. cerevisiae has found a variety of oxidation products along with taxadiene-5α-ol. It has been proposed that CYP725A2 and CYP725A1 then hydroxylate the C-13 and C-10 positions of taxadiene-5α-ol, respectively (Jennewein et al. 2001; Schoendorf et al. 2001). Furthermore, CYP725A5 and CYP725A6 catalyze the hydroxylation at the C-2 and C-7 sites of the alternative substrate taxusin (Chau and Croteau 2004; Chau et al. 2004). Further research discovered that CYP725A3 was able to introduce the hydroxy group at C-14, but it was not effective for the hydroxylation at the C-14 site in taxol (Jennewein et al. 2003) (Fig. 13). An enzyme of CYP716B subfamily from Ginkgo biloba (G. biloba) can hydroxylase the C-9 of taxane skeleton (Zhang et al. 2014).

The complex skeleton modifications in the biosynthetic pathway of taxol, especially the hydroxylation at the various sites of taxane core skeleton
Gibberellins (GAs) is an essential plant hormone that can regulate plant growth and development, such as germination, flowering, and reproduction. The simplest gibberellin GA12 is assembled by diterpene synthase (diTPS) and P450s in a tandem manner. The diterpene synthase catalyzes the formation of ent-kaurene from GGPP, followed by a sequential three-step oxidative modification by the P450 subfamily, CYP701A and CYP88A. CYP701A3 from A. thaliana acts as an ent-kaurene oxidase to catalyze the three-step continuous oxidation of the C-18 position of ent-kaurene to produce ent-kaurenoic acid. After that, CYP88A3 and CYP88A4 perform the C-7 oxidation of ent-kaurenoic acid to deliver GA12 in three consecutive oxidation steps (Helliwell et al. 1999, 2001). Also extracted from A. thaliana, CYP714A1 protein has GA12 16-carboxylation activity, while CYP714A2 can hydroxylate the C-13 or C-12 position of a variety of substrates in the metabolic pathway (Nomura et al. 2013) (Fig. 14).

Proposed partial biosynthetic pathway of gibberellins, including GA12 and GA111
Triterpene hydroxylases
Triterpenoids are higher terpenoids containing six isoprene units and most of which exist in plant resins as free states or glycosides and esters. A few triterpenoids are also found in animals. Triterpene saponins are a class of metabolites with diverse structures and biological activities from a variety of plants. 2,3-Epoxy-squalene is the common precursor to all triterpenes, which are synthesized through the cyclization by oxidosqualene cyclase (OSC). It is generally believed that the triterpene cyclization stage will form two kinds of carbocations, protosterol cation and dammarenyl cation subfamily. Protosterol cations were used as the intermediates to form tetracyclic triterpenoids, such as cycloartenol, lanosterol, protopanaxadiol, and cucurbitadienol, while the pentacyclic triterpene skeletons, like α-amyrin, β-amyrin, lupeol, and friedeleol, were derived from the dammarenyl cations (Fig. 15). Skeleton modifications catalyzed by P450s and UDP-dependent glycosylation generate a variety of sapogenins with diverse structures. The oxidation by P450s contributes a lot to these structural variants and those P450s are mainly categorized as the CYP93, CYP88, CYP716, and CYP72 families.

Triterpene skeletons derived from protosterol cation (a) and dammarenyl (b) subfamily
Ginseng is one of the most popular traditional medicinal materials and clinically display anti-stress, anticancer, anti-diabetic activities, and immune system modulation. The root contains the pharmacologically active ingredient ginsenosides derived from cytochrome P450 enzyme-catalyzed hydroxylation of the dammarenediol-II backbone and subsequent glycosyltransferase-catalyzed glycosylation. Han et al. successfully identified CYP716A47 as a protopanaxadiol synthase, which catalyzes the hydroxylation of the C-12 position of dammarenediol-II (Han et al. 2011). Subsequently, this team also isolated protopanaxadiol-6-hydroxylase CYP716A53v2 from Panax ginseng (P. ginseng) as a vital enzyme in ginsenoside biosynthesis (Han et al. 2012). CYP716A47 and CYP716A53v2 have strict substrate specificity (Fig. 16).

Proposed biosynthetic pathway for dammarenediol-type ginsenosides in P. ginseng
Mogroside is the main biologically active component extracted from Momordica grosvenori (M. grosvenori), which belongs to the cucurbitane-type tetracyclic triterpene saponins and is widely used as a natural sweetener worldwide. Mogroside is biosynthesized by a series of oxidations at the positions of C-11, C-24, and C-25 in the cucurbitadienol skeleton and subsequent glycosylations. The multifunctional enzyme CYP87D18 derived from Siraitia grosvenorii (S. grosvenorii) can carry out the C11-hydroxylation of cucurbitadienol to result in 11-hydroxycucurbitadienol, 11-oxo-cucurbitadienol, and 11-oxo-24,25-epoxy-cucurbitadienol (Zhang et al. 2016).
Many studies have revealed that P450s are clustered together with the upstream biosynthetic gene in specific terpene biosynthetic pathways. For example, 19-hydroxycucurbitadienol-25-oxidase CYP81Q58 and cucurbitadienol-19-hydroxylase CYP88L2 were found in Cucurbitaceae sativus (C. sativus) to cluster together with cucurbitadienol synthase and other P450s, providing insights for the gene mining of new P450s (Shang et al. 2014) (Fig. 17).

Proposed biosynthetic pathway for 11-oxo-cucurbitadienol and 11-oxo-24,25-epoxy-cucurbitadienol catalyzed by CYP87D18
β-Amyrin is the precursor for the biosynthesis of a series of triterpenoids, and many P450s can hydroxylate the β-amyrin skeleton to produce various triterpenoids with pharmacological activities (Fig. 18).

Biosynthesis pathways of β-amyrin skeleton triterpenoids. CYP716 subfamily in this figure: CYP716A1, CYP716A2, CYP716A12, CYP716A44, CYP716A46, CYP716A75, CYP716A94, CYP716AL1, CYP716A175, CYP716A179, CYP716A253, CYP716A52v2
Glycyrrhizin is a triterpene saponin used as a natural sweetener with pharmacological indications, such as hepatoprotective, anti-virus, and anti-allergic activities. Glycyrrhizin is biosynthesized through a series of oxidations at the C-11 and C-30 positions of β-amyrin. CYP88D6 from Glycyrrhiza uralensis (G. uralensis) catalyzes the two-step continuous hydroxylation at the C-11 position of β-amyrin to produce 11-oxo-β-amyrin (Seki et al. 2008); CYP72A154 catalyzes the three-step continuous oxidation at the C-30 position of 11-oxo-β-amyrin to form glycyrrhetinic acid (Seki et al. 2011). CYP72A63, which has a higher sequence identity with CYP72A154, can also catalyze the hydroxylation of β-amyrin at the C-30 position. In addition, Zhu et al. identified its C-30 catalytic activity toward 11-oxo-β-amyrin with a higher catalytic specificity compared with CYP72A154 (Zhu et al. 2018).
Fukushima et al. confirmed two CYP72A subfamily enzymes (CYP72A61v2 and CYP72A68v2) that are highly related to the expression of CYP93E2 and CYP716A12 in the biosynthetic pathways of hemolytic and non-hemolytic sapogenins. CYP72A61v2 catalyzes the hydroxylation at the C-22 position of 24-hydroxy-β-amyrin to produce soyasapogenol B. Moreover, CYP72A68v2 catalyzes the C-24 position by three-step continuous oxidation of oleanolic acid to release an antibacterial agent gypsogenic acid (Fukushima et al. 2013).
Shibuya et al. firstly identified a triterpene hydroxylase CYP93E1 from Glycine max (G. max) with a catalytic activity of hydroxylation at the C-24 position of β-amyrin and sophoradiol (Shibuya et al. 2006). CYP93E3 (Seki et al. 2008) and CYP93E2 identified from Medicago truncatula (M. truncatula) also have shown the β-amyrin 24-hydroxylase activity (Fukushima et al. 2011). Fukushima et al. found that CYP716A12 co-expressed with β-amyrin synthase can catalyze the hydroxylation of β-amyrin, α-amyrin, and lupeol at the C-28 position and promote the continuous oxidation to yield oleanolic acid, ursolic acid, and betulinic acid (Fukushima et al. 2011). CYP716A94 (Han et al. 2018), CYP716AL1 (Huang et al. 2012), CYP716A175 (Andre et al. 2016), CYP716A52v2 (Han et al. 2013), CYP716A179 (Tamura et al. 2017), CYP716A1, and CYP716A2 (Yasumoto et al. 2016) of the CYP716 family also have the same functions as CYP716A12, while CYP716A44 and CYP716A46 only catalyze the oxidation of β-amyrin and α-amyrin to the corresponding acids. CYP716E26 catalyzes the hydroxylation of β-amyrin at the C-6 position to generate a rare biologically active triterpene, daturadiol (Yasumoto et al. 2017). CYP716A75 and CYP716A253 only catalyze the continuous oxidation of the C-28 position of β-amyrin to oleanolic acid, while CYP716A252 catalyzes the continuous oxidation of the C-28 position of α-amyrin to ursolic acid (Misra et al. 2017). CYP716A75 was found to promote the three-step continuous hydroxylation of the C-28 position of 16α-hydroxy-β-amyrin to the precursor 16α-hydroxy-oleanolic acid for maesa saponins. CYP716Y1 identified from Bupleurum falcatum (B. falcatum) can realize the hydroxylation of α-amyrin and β-amyrin at the C-16 position (Moses et al. 2014). Alternatively, CYP72A397 carries out the C-23 hydroxylation of oleanolic acid to give hederagenin with intriguing pharmacological properties (Han et al. 2018).
Tzin et al. reported other two CYP72A subfamily enzymes. CYP72A67 was found to catalyze the hydroxylation at the C-2 position of oleanolic acid and hederagenin. CYP72A68 catalyzes the three-step continuous oxidation of oleanolic acid to afford an antibacterial agent gypsogenic acid (Tzin et al. 2019). At present, a variety of P450 enzymes using β-amyrin as substrate have been identified in dicots, but only CYP51H10 has been discovered in monocots (Qi et al. 2006). CYP51H10 from Avena strigosa (A. strigosa) is a multifunctional P450 that can catalyze the oxidation of the C ring and D ring of β-amyrin to furnish the important precursor 12,13-β-epoxy-16β-hydroxy-β-amyrin for avenacins (Geisler et al. 2013).
Tetraterpene hydroxylases
Tetraterpenoids feature a complex structure composed of eight isoprene units, of which carotenoids are the most representative tetraterpene pigments that are widely found in photosynthetic bacteria, some archaea, fungi, plants, and animals. As a photoprotective agent, antioxidant, and precursor of plant hormones, carotenoids are widely present in plants. Animals cannot synthesize carotenoids de novo, so those found in animals are usually ingested from plants and partly modified through metabolic reactions, rendering the structural diversity of animal carotenoids.
There exist two kinds of carotenoids: carotenes, composed of hydrocarbons, including α-carotene, β-carotene, and γ-carotene; the other is xanthophylls containing several oxygen atoms which are introduced by various hydroxylases, such as hydroxyl, carboxyl, aldehyde, and epoxidation (Maoka 2020) (Fig. 19).

a Basic structures of carotenoids and end groups; b structures of typical carotenes and xanthophylls
Two classes of structurally unrelated hydroxylases that can modify the rings of carotenes to xanthophylls, one is the heme-containing P450s and the other is the non-heme di-iron hydroxylases. CYP175A1, derived from the thermostable bacterium Thermus thermophilus HB27, shows a β-ring hydroxylase activity in the heterologous host E. coli, which was identified as the first β-carotene hydroxylase in the P450 superfamily.
So far, all the cytochrome P450‐type carotene hydroxylases identified from plants belong to the CYP97 family. CYP97A3 and CYP97C1 from the phenotypes of the knock-out mutants of A. thaliana have exhibited activities against carotenoids. CYP97A3 can hydroxylate the two β-rings of β-carotene to produce zeaxanthin (Kim and DellaPenna 2006), while CYP97C1 can hydroxylate the β-ring and ε-ring to produce lutein (Tian and DellaPenna 2004). CYP97A4 and CYP97C2 from Oryza sativa have similar functions like the above two enzymes. CYP97A4 mainly acts as a β-ring carotene hydroxylase with minor activity toward the ε-rings, while CYP97C2 specifies the conversion of the ε-rings. Combined with the distribution of these hydroxylases and the accumulation of metabolites in plants, it is considered that Clan A enzymes in the CYP97 family are putative β-rings carotene hydroxylases, while Clan C enzymes are potential ε-rings carotene hydroxylases (Quinlan et al. 2007). Members of the CYP97B subfamily are more distant from Clans A and C with a wider distribution in plants. The first carotene β-rings hydroxylase CYP97B29 identified in Red algae can hydroxylate β-carotene to yield zeaxanthin (β,β-carotene-3,3′-diol). This hydroxylation reaction involves the extraction of hydrogen and the hydroxylation of the C-3 positions of the two β-rings (Yang et al. 2014). In addition, CYP97B3 was also found to be associated with carotenoid metabolism in A. thaliana (Kim et al. 2010) (Fig. 20).

Biosynthetic pathway of Xanthophylls catalyzed by P450‐type carotene hydroxylases. LYCB lycopene β-cyclase, LYCE lycopene ε-cyclase
Two non-heme di-iron hydroxylases CHXB (β-ring carotene hydroxylase) and CHXE (ε-ring carotene hydroxylase) from Lycium chinense (L. chinense) can hydroxylate α-carotene and β-carotene to form lutein (Zhao et al. 2014). In addition, a series of non-heme carotenoid hydroxylases from A. thaliana, green algae, and other plants, as well as from photosynthetic and non-photosynthetic bacteria have been cloned and functionally characterized in heterologous hosts (Galpaz et al. 2006; Kim et al. 2009; Linden 1999; Qi et al. 2020; Tian and DellaPenna 2004).
The phylogenetic relationships of terpene hydroxylases
CYP450s involved in this article and their corresponding functions are listed in Table 1. As described above, on one hand, hydroxylases from different families or even different species may be able to accept terpene skeleton of a similar structure. For example, taxane hydroxylases derived from Taxus sp. all belong to CYP725A subfamily; however, an enzyme belonging to CYP716B subfamily derived from G. biloba was found to be able to hydroxylate the C-9 position of the taxane skeleton. Similarly, enzymes of the CYP97 family are mainly involved in the biosynthesis of tetrapenoid zeaxanthin; however, CYP175A1 was also found to hydroxylate β-cryptoxanthin to produce zeaxanthin. On the other hand, the metabolic pathway of some structurally similar substrates often involves enzyme from several specific families. For example, the vast majority of biocatalysts capable of hydroxylating or oxidizing β-amyrin backbone-type triterpenes belong to CYP716, CYP88, and CYP72 (Fig. 18). It seems that some associations exist between the specific CYP450 family and their substrate spectrum.
We established the phylogenetic relationship of a total of 108 terpene hydroxylases involved in this paper, with the purpose to explore the connections between these different hydroxylases from the perspective of evolution (Fig. 21). In general, CYP76 and CYP71 families are involved in the biosynthesis of monoterpenes (Fig. 2); CYP71, CYP264, and CYP260 families are involved in the biosynthesis of sesquiterpenes (Fig. 8); CYP726, CYP725, and CYP716 families are mainly involved in the biosynthesis of diterpenes; and CYP97 and CYP175 are involved in the biosynthesis of tetraterpene (Fig. 20). It is obvious that as the size of substrate backbone gradually increases, the location of the corresponding CYP450 families on the phylogenetic tree gradually transits from one side (feature with blue and green) to another side (feature with red and orange). Some individual exceptions exist, for example, CYP701A3 is able to hydroxylate the C-18 position of GA (Fig. 14), yet it is more closely related to the CYP71 and CYP76 families and more distant from the branch where other triterpene hydroxylases are located. However, individual examples did not hinder the overall phylogenetic trend.

The phylogenetic tree of CYP450s involved in this article
Different CYP450s capable to accept similar substrates often have a close phylogenetic relationship. Besides, it is obvious from the evolutionary tree that biocatalysts of these three families (CYP76, CYP71, and CYP726) evolved from the same ancestral enzyme. As a result, it is presumed there exists a close evolutionary connection between monoterpenes hydroxylases and sesquiterpenes hydroxylases. As a contrast, the core skeletons of diterpenes, triterpenes, and tetrapenes are often rather large, and it seems that their corresponding hydroxylases are more evolved from another ancestral enzyme.
These phenomena provide some novel ideas for the digging of new terpene hydroxylases. When we need to search for hydroxylases with specific substrate backbones, it is feasible to mine and screen CYP450s from some specific families. For instance, it seems that there exists a higher probability of finding new monoterpene hydroxylases from CYP76, CYP71 families, or other families which have a closer phylogenetic relationship with these two CYP450 families.
Optimize expression, activity, and selectivity of terpenoid hydroxylases
For a long time, terpenoids with intriguing physiological and pharmacological activities have emerged as the appealing topics in the community. From the extraction of active ingredients to the isolation and purification of single compound, more and more terpenoids have been characterized and investigated in the biological pathways.
CYP450s involved in the biosynthetic pathway of terpenoids remain a main bottleneck for clarifying the detailed function, retarding to display the full capacity of oxidases in producing significant substances. These P450s have problems, such as low expression, poor stability, as well as low activity. At the same time, the catalytic promiscuity, which means these hydroxylases can accept a variety of substrates or catalyze one substrate to generate a variety of products, is also a significant obstacle resulting in the dispersion of metabolic pathways and further reducing the yield of desired products. At present, there are many studies focusing on solving these problems, among which the heterologous expression of these hydroxylases has been significantly improved. Meanwhile, some progresses have been made aiming at the catalytic promiscuity and low activity of these hydroxylases. The challenges and corresponding solutions of these CYP450s are summarized in the following part.
Optimize the functional expression of terpene hydroxylases
Synthetic biology techniques generally choose to construct metabolic pathways for the terpenoid production in easily cultured organisms, such as the bacterial system E. coli or the fungal system S. cerevisiae. Most terpene hydroxylases, especially those derived from eukaryotes, belong to type II P450s and bind to the endoplasmic reticulum membrane. S. cerevisiae are generally more suitable for the expression of CYP450s of eukaryotic origin because of their natural endosomal system. Optimizing promoter strength as well as increasing gene copy number are methods that are usually used to improve protein expression in S. cerevisiae (Walls et al. 2021). However, in general, the functional expression of these CYP450s in E. coli is challenging because of the inherent limitations of the bacterial platform. Firstly, E. coli lacks membrane structures, such as endoplasmic reticulum, which is incompatible with the translation of the membrane signaling modules of membrane-bound CYP450s. Secondly, the lack of CYP450 reductases (CPRs) in the prokaryotic system cannot transmit electrons for CYP, that hinders the catalytic function of CYP. Therefore, in general, membrane proteins, such as CYP725A4, are difficult to functionally express in bacterial systems.
A variety of measures have been developed to achieve the functional expression of these membrane-bound CYP450s in easy-to-handle model heterologous host E. coli, including N-terminal modification or co-expression of chaperones.
N-terminal modification
The N-terminus of type II P450s are often composed of hydrophobic amino acids which can bind to the endoplasmic reticulum and exert catalytic function. The predict protein software (http://www.predictprotein.org/) can be used to identify the transmembrane region of these membrane-bound P450s. This hydrophobic sequence always becomes an obstacle to heterologous expression in E. coli. As a result, part of the N-terminal hydrophobic sequence was often truncated to increase the solubility of the protein and promote its correct folding. On the contrary, some hydrophilic sequences can be artificially added in N-terminus.
The inclusion body of 5α-hydroxylase for taxadiene (CYP725A4 from Taxus) is obtained by directly expressing the enzyme in E. coli, and several groups have developed various modification strategies to promote its soluble expression. Rouck et al. obtained three constructs MA, 2b1, and 17α through different modifications to the N-terminal of CYP725A4 (Rouck et al. 2017). MA herein represents that the N-terminal 59 amino acids of CYP725A4 have been truncated and Met-Ala was added to the starting position which can help increase the soluble expression; 2b1 is based on the truncation of 59 amino acids at the N-terminal and replaced with a peptide MAKKTSSKGKLPPGPS from the N-terminal of rabbit CYP2b1; 17α means that the N-terminal 24 amino acids of CYP725A4 are truncated and a sequence of MALLLAVF from bovine steroid hydroxylase is added. The yields of three purified proteins obtained in E. coli were 1.54 nmol/L, 1.76 nmol/L and 4.83 nmol/L, respectively, indicating that this N-terminal modification method has a certain positive effect on the soluble expression of P450s.
Ajikumar et al. truncated the N-terminus of CYP725A4 and CPR with different lengths through transmembrane engineering by adding MALLLAVF into the N-terminus of CYP725A4. The two truncated proteins were connected through the GSTGS linker to construct a chimeric protein (Fig. 22). Among the constructed chimeric proteins, At24T5αOH-tTCPR has the highest conversion rate to the substrate taxadiene, of which CYP725A4 was truncated by 24 amino acids and Taxus-derived CPR truncated 74 amino acids in the N-terminus endoplasmic reticulum binding region (Ajikumar et al. 2010).

Schematic diagram of chimeric protein construction. At24T5αOH-tTCPR was taken as a typical example
Co-expression of molecular chaperones
Some P450s co-expressed with molecular chaperones to provide a suitable hydrophilic environment to help the correct folding of P450s and the proper insertion of heme. Ichinose et al. used different modification methods to enable 304 P450s derived from fungi to be functionally expressed in E. coli. Among them, CYP505D8v1, CYP5137A4v1, and CYP505D6 use molecular chaperones GroES/GroEL to help proteins fold correctly, and their volume titers reached 1333 nmol/L, 1820 nmol/L, and 665 nmol/L, respectively. CYP5139D7v1, CYP5147B1, and CYP5150A2 were truncated to varying degrees of N-terminal, and the co-expression of GroES/GroEL also helped these P450s fold correctly. CYP5037E1 and CYP5149A1 were reconstructed by N-terminal modification and replaced with the N-terminal domain of CYP5144C1 to construct a chimeric protein to achieve a volume titer of 2000 nmol/L, which is currently the highest yield of fungal P450s expressed in E. coli. These reports have important references for expressing P450s from other eukaryotic sources in E. coli (Fatima et al. 2021; Ichinose et al. 2015).
Improve the catalytic activity and chemo- or regioselectivity of terpene hydroxylases
Co-expression of external redox partner
Cytochrome P450 reductase (CPR), also known as NADPH-dependent cytochrome P450 oxidoreductase, is a membrane-bound oxidoreductase embedded in the cytosolic side of the endoplasmic reticulum. CPR is indispensable in most CYP450-catalyzed reactions, responsible for sequentially delivering two electrons from the cofactor NAD(P)H to the heme Fe. The first electron in CPR reduces the heme Fe and promotes the O2 binding to form a superoxide complex (3), and the second electron further reduces 3 to a peroxide complex (Fe3+OO)2− P450 (4). The subsequent reaction oxidizes the substrate and produces a molecule of water (Fig. 23). CPR modulates the activities of P450s through altering the electron transfer processes. In addition, the protein–protein interaction (PPI) between chimera CYP450 and CPR may alter the spatial conformation of CYP450, thus affecting the substrate binding (Zhang et al. 2022; Sun et al. 2020).

Typical catalytic cycle of P450s for hydroxylations. The exact structure of heme is marked on this diagram
In general, the electron transfer system expression hosts, such as E. coli and S. cerevisiae, are not completely matched to P450s originated from plants. Therefore, it is often necessary to co-express homologous CPR helping P450s to exert their physiological effects. At present, the most commonly used eukaryotic expression hosts S. cerevisiae WAT11 and WAT21 are constructed by using the CPRs from A. thaliana-derived ATR1 and ATR2 to replace the endogenous genes of CPR1 and CPR2, respectively (Pompon et al. 1996).
Zhu et al. screened six CPRs derived from different plants, including ATR1 and ATR2 from A. thaliana, MTR2 and MTR3 from Medicago sativa, LjCPR1 from Lotus japonicus, and GuCPR1 from G. uralensis. Their coupling efficiencies with CYP72A154, a CYP450 involved in the biosynthesis of glycyrrhetinic acid, were examined by inserting the genes of CYP72A154 and the corresponding CPR into the genome of engineered S. cerevisiae INVSc1. The production of GA and other related triterpenoids were varied significantly when different CPR was used, with the engineered strain containing GuCPR1 had the highest titer of GA. This phenomenon revealed that CPRs have the capability to affect the catalytic activity of CYP450 and further altered the overall productivity of terpenoids (Zhu et al. 2018).
Chang et al. showed when CYP706B1 (cadinene C-8 hydroxylase) was co-transformed into the engineered E. coli with aaCPRct from Artemisia annua with a 1.8-fold decrease in the overall productivity of 8-hydroxycadinene, compared with that co-transformed with ctCPR from Candida tropicalis. Since these two CPRs did not show apparent difference in the in vitro cytochrome c reduction activity, this phenomenon seems to be attributed to the protein–protein interactions between CYP450 and CPRs (Chang et al. 2007).
The low electron transfer efficiency between CPR and P450 is one of the main reasons rendering the low catalytic activity of P450s. Very recently, Park et al. connected CPR and P450 with the self-assembled scaffold protein CipB from Photorhabdus luminescens, respectively. After connected with this scaffold protein, CPR and P450 can spontaneously form the protein crystal inclusion (PCI) in E. coli, which shortened the distance between CPR and P450, as a result increasing the efficiency of CPR transferring electrons to P450. This strategy was further applied to the P450s related to the biosynthesis pathway of lutein, (+)-nootkatone, apigenin, and l-3,4-dihydroxyphenylalanine (l-DOPA), respectively. The production of these compounds in engineered E. coli was significantly increased, proving that using scaffold proteins to assist electron transfer between CPRs and P450s was capable for universally application (Park et al. 2022).
Additionally, a single CPR can supply electrons to multiple P450s in natural product biosynthetic pathways. It was founded that the ratio of CPR and CYP450s achieve 1:15 in the microsomal membrane (Jensen and Møller 2010). To minimize the generation of toxic reactive oxygen species caused by the disruption of the degree of coupling between the CPR and CYP450s, Walls et al. expressed CYP725A4 under GAL1 promoter to maximize the expression of CYP450, while expressed CPR under the weak GAL3 promoter in S. cerevisiae (Walls et al. 2021).
Molecular modification of terpene hydroxylases
Eukaryotic terpenoid hydroxylases, especially those derived from plants, are typically poorly expressed in heterologous hosts with low stability. These membrane proteins are difficult to be purified or crystallized, and as a result, structural information about these enzymes are extremely rare. On the other hand, lacking of high-throughput screening methods render traditional directed evolution methods, such as error-prone PCR, which require the screening of a large number of mutant libraries, to be difficult to apply in the modification of terpene hydroxylases.
Much of what is known about CYP450 structure–function relationships and molecular modification of the activity and selectivity of terpene hydroxylases is therefore based on the homologous models of the structure of CYP450s already clarified from bacterium. Small mutant libraries are designed toward the active centers or a specific secondary structure area of those hydroxylases, and the viability, regional selectivity, and stereoselectivity of mutants were further characterized by intracellular fermentation experiments. Additionally, hydroxylases capable of hydroxylating different locations of the same terpene backbone often originate from a same CYP450 subfamily with high sequence identity among each other. Therefore, analysis of the evolutionary data of amino acid is also a feasible method to locate the key amino acids that affect the selectivity or activity of terpenoid hydroxylases.
-
(1)
Manipulate the regioselectivity of terpenoid hydroxylases
Limonene-3-hydroxylases CYP71D13 and CYP71D15 and limonene-6-hydroxylase CYP71D18 have been identified from mint oil gland cDNA libraries. CYP71D13 and CYP71D15 were found to exhibit 93% sequence identity with each other and showed around 70% sequence identity with CYP71D18. The exploration of regioselectivity of limonene hydroxylases mainly started from the sequence alignment results. Firstly, through building chimeric enzymes to exchange large restriction fragments and exchange of subdomains of CYP71D15 and CYP71D18, a subdomain with a total length around 20 amino acids is localized, which had a direct effect on the hydroxylation regiospecificity of the product. It was found that only 5 amino acids were different in this subdomain, and therefore, reciprocal site-directed mutagenesis was designed for these five mutation sites. Interestingly, after changing Phe363 of CYP71D18 to Ile, the selectivity of its hydroxylation position was reversed from the C-6 to C-3 position, indicating that the residues at site 363 play an important role in the binding of the substrate and affected the regiospecific preference (Schalk and Croteau 2000).
Moreover, Mau et al. identified CYP71D174 from Perilla frutescens as limonene-7-hydroxylase by using the cDNA sequence of limonene-3-hydroxylase and limonene-6-hydroxylase as probes. CYP71D174 is 63% and about 69% identical with CYP71D18 and CYP71D13, respectively. The amino acid of Phe at position 363 of CYP71D174 catalyzes (−)-limonene to produce the C-7, C-3, and C-6 oxygenated products. This phenomenon showed that besides the amino acid at position 363, there exist other amino acids involved in the immobilization of the substrate in the active center (Mau et al. 2010).
-
(2)
Expanding the substrate spectrum and modify substrate heterogeneity of terpenoid hydroxylases
In the biosynthesis of cucumber cucurbitacin C (CuC), CYP87D20 from Cucumis sativus L. catalyzes the C11-oxidation of the cucurbitacin skeleton to produce 11-carbonyl-cucurbitadienol (Zhou et al. 2016). Based on the crystal structure of CYP51 derived from S. cerevisiae, Li et al. constructed a homologous model of CYP87D20 with Rosetta and further carried out three rounds of structural and data-driven direct evolutions to modify CYP87D20 (formation of cucurbitadienol-C11 ketone and C20 hydroxyl) into an efficient cucurbitadienol-C11-hydroxylase (Fig. 24).
Fig. 24 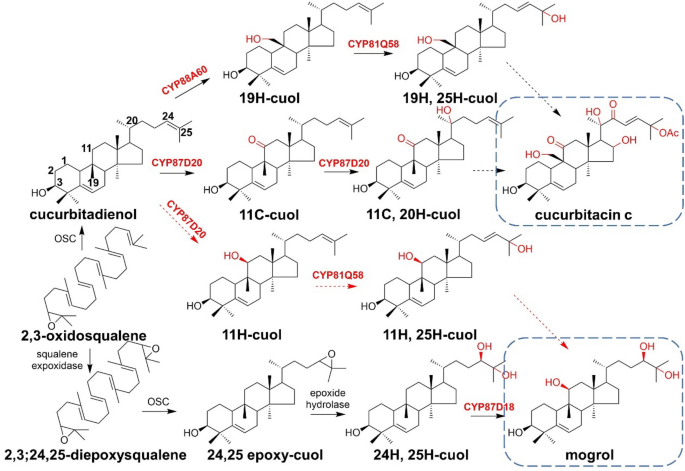
Biosynthesis pathways of CuC and mogrol. Proposed metabolic flux to mogrol is indicated with red arrow. OSC oxidosqualene cyclase, cuol cucurbitadienol
In the first round of engineering (R1), amino acids within the range of substrate 6 Å that may affect the specificity of the substrate/product were analyzed. Some mutants produce an increased amount of 11-hydroxyl-cucurbitadienol, as well as a decreased amount of 11-carbonyl-cucurbitadienol or 11-carbonyl-20-hydroxyl-cucurbitadienol. In the second round of engineering (R2), the amino acids around the positive mutants screened by R1 were selected, which further improved the selectivity and activity. In the third round of mutations, 7 mutants from R1 and R2 were selected, and 2 to 3 mutation sites were randomly inserted into these 7 mutants by computer algorithm, and finally more than 27,000 potential mutants were obtained in silico and detected by experiment. After three rounds of modifications and evaluations, Li et al. successfully carries out a single specific hydroxylation at the C-11 position for CYP87D20, thereby increasing the metabolic flux to mogrol (Li et al. 2019).
-
(3)
Manipulate the chemo- and regioselectivity of terpenoid hydroxylases
Most plant-derived CYP450s can act on various substrates to generate a spectrum of products. Enzymatic multi-specificity and promiscuity are entirely new propositions in metabolic engineering, and canonical enzyme engineering strategies for modulating the activity of enzymes are largely inapplicable for addressing this phenomenon (Yadav 2014).
CYP725A4, the first hydroxylase involved in the biosynthetic pathway of taxol, catalyzing taxadiene to form multiple products, including several mono- and di-hydroxylated diterpenoids. Through homology modeling and molecular docking, Edgar et al. selected amino acids within the activation center to construct a mutant library with the aim to improve the yield of di-hydroxylated product. Although most of the mutants did not achieve enhanced selectivity, T380S improved the percentage of 5(12)-oxa-3(11)-cyclo-taxan-10-ol in a variety of products. The presumed production process of this compound is shown in Fig. 25 (Edgar et al. 2017).
Fig. 25 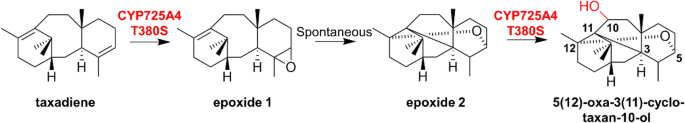
Proposed pathway from taxadiene to 5(12)-oxa-3(11)-cyclo-taxan-10-ol
CYP72A63 derived from M. truncatula has the capability of oxidizing 11-oxo-β-amyrin to produce a range of rare licorice triterpenoids with a low selectivity, including three C-30 oxidized products glycyrrhetol, glycyrrhetaldehyde, and glycyrrhetinic acid, and a C-29 hydroxynated product 29-OH-11-oxo-β-amyrin. Sun et al. identified the key amino acids affecting the specificity of CYP72A63 through homologous modeling and molecular docking. The obtained mutations can specifically convert 11-oxo-β-amyrin to the corresponding hydroxyl, aldehyde, and carboxylic acid, respectively (Fig. 26).
Fig. 26 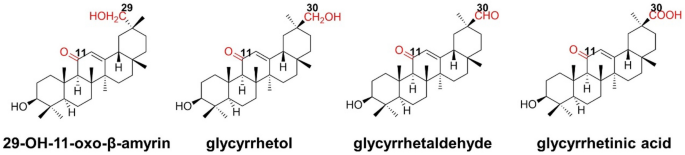
Four kinds of triterpenoids produced by CYP72A63 with 11-oxo-β-amyrin as substrate
Firstly, substrate 11-oxo-β-amyrin was docked into the active center of CYP72A63. It was found that C-30 and C-29 were all within the catalytic range of heme Fe, while C-30 was closer, which was consistent with the higher oxidation preference at the site of C-30. Secondly, to exploit the consecutive three-step oxidation at C-30, glycyrrhetol was docked with CYP72A63 to reveal a less fitness than 11-oxo-β-amyrin in CYP72A63. Based on the model, it is thus speculated that the hydrophilic hydroxy group at the C-30 position of glycyrrhetol is difficult to enter the active center because of the hydrophobic force of the active site enhanced by the methyl side chain of Thr338.
After mutating Thr338 to Ser, the side-chain hydroxyl group of Ser points to the active center, increasing the hydrophilicity of the active pocket and enabling glycyrrhetol to be almost completely converted to glycyrrhetinic acid. At the same time, this mutation also makes C-29 farther away from the heme Fe, avoiding the formation of byproduct 29-OH-11-oxo-β-amyrin. Additionally, when Leu509 located in the distal active pocket of CYP72A63 is mutated to Ile, Thr, and Asn, that can significantly hinder the further oxidation of glycyrrhetol, thereby generating glycyrrhetol in high yield (Fig. 27). These mutants constructed based on the homologous models enabled the generation of different licorice-type triterpenoids with high regional and chemical selectivity (Sun et al. 2020).
Fig. 27 
a The structure model of CYP72A63 with 11-oxo-β-amyrin; b The structure model of CYP72A63 (T338S) with glycyrrhetol




Summary and outlook
Biocatalytic oxidation equips enormous potential in effectively obtaining bioactive and medicinally relevant terpenoids. Especially, the site-specific and stereo-specific hydroxylation mainly mediated by cytochrome P450 monooxygenases addressed lots of unsolved challenges in total synthesis of natural products by now. A series of monoterpene, sesquiterpene, diterpene, and other terpene hydroxylases have been discovered in plants, animals, and microorganisms through genome mining and cDNA library screening based on homologous sequences, which has made an important contribution to the study of terpenoids biosynthesis.
Additionally, the number of enzymes that have been identified are still relatively limited compared with the natural resources. With advances in sequencing technology, it is anticipated that more new hydroxylases would be discovered and characterized. These hydroxylases usually have low heterologous expression, poor stability, and low catalytic efficiency. Through N-terminal modification and redox partner optimization, the expression of P450s in prokaryotic host had been increased apparently. With the development of new technology like cell-free expression system (Walter et al. 2022), it is expected that expression no longer being the bottleneck restricting the further investigation towards terpene hydroxylases.
Finally, most eukaryotic terpene hydroxylase belongs to type II CYP450s, their activity and expression all restricted by the inherent structure and catalytic properties. In view of this, it is also a feasible method to explore and modify other oxygenase superfamilies (Renata 2021; Zwick and Renata 2020), so that they can catalyze corresponding terpene substrates to generate more valuable terpenoids and structural derivatives. Researches on hydroxylases involved in terpenoid biosynthesis is of great significance for promoting the discovery and synthesis of more bioactive medicinal resources.
Availability of data and materials
Not applicable.
References
Ajikumar PK, Xiao WH, Tyo KE, Wang Y, Simeon F, Leonard E, Mucha O, Phon TH, Pfeifer B, Stephanopoulos G (2010) Isoprenoid pathway optimization for taxol precursor overproduction in Escherichia coli. Science 330:70–74
Andre CM, Legay S, Deleruelle A, Nieuwenhuizen N, Punter M, Brendolise C, Cooney JM, Lateur M, Hausman JF, Larondelle Y, Laing WA (2016) Multifunctional oxidosqualene cyclases and cytochrome P450 involved in the biosynthesis of apple fruit triterpenic acids. New Phytol 211:1279–1294
Behrendorff JB, Gillam EM (2017) Prospects for applying synthetic biology to toxicology: future opportunities and current limitations for the repurposing of cytochrome P450 systems. Chem Res Toxicol 30:453–468
Besseau S, Kellner F, Lanoue A, Thamm AM, Salim V, Schneider B, Geu-Flores F, Hofer R, Guirimand G, Guihur A, Oudin A, Glevarec G, Foureau E, Papon N, Clastre M, Giglioli-Guivarc’h N, St-Pierre B, Werck-Reichhart D, Burlat V, De Luca V, O’Connor SE, Courdavault V (2013) A pair of tabersonine 16-hydroxylases initiates the synthesis of vindoline in an organ-dependent manner in Catharanthus roseus. Plant Physiol 163:1792–1803
Boutanaev AM, Moses T, Zi J, Nelson DR, Mugford ST, Peters RJ, Osbourn A (2015) Investigation of terpene diversification across multiple sequenced plant genomes. Proc Natl Acad Sci USA 112:E81–E88
Bozic D, Papaefthimiou D, Bruckner K, de Vos RC, Tsoleridis CA, Katsarou D, Papanikolaou A, Pateraki I, Chatzopoulou FM, Dimitriadou E, Kostas S, Manzano D, Scheler U, Ferrer A, Tissier A, Makris AM, Kampranis SC, Kanellis AK (2015) Towards elucidating carnosic acid biosynthesis in Lamiaceae: functional characterization of the three first steps of the pathway in Salvia fruticosa and Rosmarinus officinalis. PLoS ONE 10:e0124106
Cankar K, van Houwelingen A, Bosch D, Sonke T, Bouwmeester H, Beekwilder J (2011) A chicory cytochrome P450 mono-oxygenase CYP71AV8 for the oxidation of (+)-valencene. FEBS Lett 585:178–182
Chacon-Morales PA (2022) Unprecedented diterpene skeletons isolated from vascular plants in the last twenty years (2001–2021). Phytochemistry 204:113425
Chang MCY, Eachus RA, Trieu W, Ro DK, Keasling JD (2007) Engineering Escherichia coli for production of functionalized terpenoids using plant P450s. Nat Chem Biol 3:274–277
Chau M, Croteau RB (2004) Molecular cloning and characterization of a cytochrome P450 taxoid 2alpha-hydroxylase involved in taxol biosynthesis. Arch Biochem Biophys 427:48–57
Chau M, Jennewein S, Walker K, Croteau RB (2004) Taxol biosynthesis: molecular cloning and characterization of a cytochrome P450 taxoid 7 beta-hydroxylase. Chem Biol 11:663–672
Chen CC, Malwal SR, Han X, Liu W, Ma L, Zhai C, Dai L, Huang JW, Shillo A, Desai J, Ma X, Zhang Y, Guo RT, Oldfield E (2020) Terpene cyclases and prenyltransferases: structures and mechanisms of action. ACS Catal 11:290–303
Chien TY, Huang SK, Lee CJ, Tsai PW, Wang CC (2016) Antinociceptive and anti-inflammatory effects of zerumbone against mono-iodoacetate-induced arthritis. Int J Mol Sci 17:249
Chiu CC, Keeling CI, Bohlmann J (2019a) The cytochrome P450 CYP6DE1 catalyzes the conversion of alpha-pinene into the mountain pine beetle aggregation pheromone trans-verbenol. Sci Rep 9:1477
Chiu CC, Keeling CI, Henderson HM, Bohlmann J (2019b) Functions of mountain pine beetle cytochromes P450 CYP6DJ1, CYP6BW1 and CYP6BW3 in the oxidation of pine monoterpenes and diterpene resin acids. PLoS ONE 14:e0216753
Corinna W, Henrik TS (2013) Cytochrome P450-enzymes involved in the biosynthesis of mono- and sesquiterpenes. Phytochem Rev 14:7–24
Edgar S, Li FS, Qiao KJ, Weng JK, Stephanopoulos G (2017) Engineering of taxadiene synthase for improved selectivity and yield of a key taxol biosynthetic intermediate. ACS Synth Biol 6:201–205
Eljounaidi K, Cankar K, Comino C, Moglia A, Hehn A, Bourgaud F, Bouwmeester H, Menin B, Lanteri S, Beekwilder J (2014) Cytochrome P450s from Cynara cardunculus L. CYP71AV9 and CYP71BL5, catalyze distinct hydroxylations in the sesquiterpene lactone biosynthetic pathway. Plant Sci 223:59–68
Fatima K, Naqvi F, Younas H (2021) A review: molecular chaperone-mediated folding, unfolding and disaggregation of expressed recombinant proteins. Cell Biochem Biophys 79:153–174
Fu N, Yang ZL, Pauchet Y, Paetz C, Brandt W, Boland W, Burse A (2019) A cytochrome P450 from the mustard leaf beetles hydroxylates geraniol, a key step in iridoid biosynthesis. Insect Biochem Mol Biol 113:103212
Fukushima EO, Seki H, Ohyama K, Ono E, Umemoto N, Mizutani M, Saito K, Muranaka T (2011) CYP716A subfamily members are multifunctional oxidases in triterpenoid biosynthesis. Plant Cell Physiol 52:2050–2061
Fukushima EO, Seki H, Sawai S, Suzuki M, Ohyama K, Saito K, Muranaka T (2013) Combinatorial biosynthesis of legume natural and rare triterpenoids in engineered yeast. Plant Cell Physiol 54:740–749
Galpaz N, Ronen G, Khalfa Z, Zamir D, Hirschberg J (2006) A chromoplast-specific carotenoid biosynthesis pathway is revealed by cloning of the tomato white-flower locus. Plant Cell 18:1947–1960
Gavira C, Hofer R, Lesot A, Lambert F, Zucca J, Werck-Reichhart D (2013) Challenges and pitfalls of P450-dependent (+)-valencene bioconversion by Saccharomyces cerevisiae. Metab Eng 18:25–35
Geisler K, Hughes RK, Sainsbury F, Lomonossoff GP, Rejzek M, Fairhurst S, Olsen CE, Motawia MS, Melton RE, Hemmings AM, Bak S, Osbourn A (2013) Biochemical analysis of a multifunctional cytochrome P450 (CYP51) enzyme required for synthesis of antimicrobial triterpenes in plants. Proc Natl Acad Sci USA 110:e3360-3367
Gesell A, Blaukopf M, Madilao L, Yuen MM, Withers SG, Mattsson J, Russell JH, Bohlmann J (2015) The gymnosperm cytochrome P450 CYP750B1 catalyzes stereospecific monoterpene hydroxylation of (+)-sabinene in thujone biosynthesis in western redcedar. Plant Physiol 168:94–106
Giddings LA, Liscombe DK, Hamilton JP, Childs KL, DellaPenna D, Buell CR, O’Connor SE (2011) A stereoselective hydroxylation step of alkaloid biosynthesis by a unique cytochrome P450 in Catharanthus roseus. J Biol Chem 286:16751–16757
Ginglinger JF, Boachon B, Hofer R, Paetz C, Kollner TG, Miesch L, Lugan R, Baltenweck R, Mutterer J, Ullmann P, Beran F, Claudel P, Verstappen F, Fischer MJ, Karst F, Bouwmeester H, Miesch M, Schneider B, Gershenzon J, Ehlting J, Werck-Reichhart D (2013) Gene coexpression analysis reveals complex metabolism of the monoterpene alcohol linalool in Arabidopsis flowers. Plant Cell 25:4640–4657
Girhard M, Klaus T, Khatri Y, Bernhardt R, Urlacher VB (2010) Characterization of the versatile monooxygenase CYP109B1 from Bacillus subtilis. Appl Microbiol Biotechnol 87:595–607
Guo J, Ma X, Cai Y, Ma Y, Zhan Z, Zhou YJ, Liu W, Guan M, Yang J, Cui G, Kang L, Yang L, Shen Y, Tang J, Lin H, Ma X, Jin B, Liu Z, Peters RJ, Zhao ZK, Huang L (2016) Cytochrome P450 promiscuity leads to a bifurcating biosynthetic pathway for tanshinones. New Phytol 210:525–534
Hamberger B, Ohnishi T, Hamberger B, Seguin A, Bohlmann J (2011) Evolution of diterpene metabolism: sitka spruce CYP720B4 catalyzes multiple oxidations in resin acid biosynthesis of conifer defense against insects. Plant Physiol 157:1677–1695
Han JY, Kim HJ, Kwon YS, Choi YE (2011) The Cyt P450 enzyme CYP716A47 catalyzes the formation of protopanaxadiol from dammarenediol-II during ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol 52:2062–2073
Han JY, Hwang HS, Choi SW, Kim HJ, Choi YE (2012) Cytochrome P450 CYP716A53v2 catalyzes the formation of protopanaxatriol from protopanaxadiol during ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol 53:1535–1545
Han JY, Kim MJ, Ban YW, Hwang HS, Choi YE (2013) The involvement of beta-amyrin 28-oxidase (CYP716A52v2) in oleanane-type ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol 54:2034–2046
Han JY, Chun JH, Oh SA, Park SB, Hwang HS, Lee H, Choi YE (2018) Transcriptomic analysis of kalopanax septemlobus and characterization of KsBAS, CYP716A94 and CYP72A397 genes involved in hederagenin saponin biosynthesis. Plant Cell Physiol 59:319–330
Helliwell CA, Poole A, James Peacock W, Dennis ES (1999) Arabidopsis ent-Kaurene oxidase catalyzes three steps of gibberellin biosynthesis. Plant Physiol 119:507–510
Helliwell CA, Chandler PM, Poole A, Dennis ES, Peacock WJ (2001) The CYP88A cytochrome P450, ent-kaurenoic acid oxidase, catalyzes three steps of the gibberellin biosynthesis pathway. Proc Natl Acad Sci USA 98:2065–2070
Hofer R, Boachon B, Renault H, Gavira C, Miesch L, Iglesias J, Ginglinger JF, Allouche L, Miesch M, Grec S, Larbat R, Werck-Reichhart D (2014) Dual function of the cytochrome P450 CYP76 family from Arabidopsis thaliana in the metabolism of monoterpenols and phenylurea herbicides. Plant Physiol 166:1149–1161
Huang L, Li J, Ye H, Li C, Wang H, Liu B, Zhang Y (2012) Molecular characterization of the pentacyclic triterpenoid biosynthetic pathway in Catharanthus roseus. Planta 236:1571–1581
Huang ZY, Ye RY, Yu HL, Li AT, Xu JH (2021) Mining methods and typical structural mechanisms of terpene cyclases. Bioresour Bioprocess 8:66
Ichinose H, Hatakeyama M, Yamauchi Y (2015) Sequence modifications and heterologous expression of eukaryotic cytochromes P450 in Escherichia coli. J Biosci Bioeng 120:268–274
Ikezawa N, Gopfert JC, Nguyen DT, Kim SU, O’Maille PE, Spring O, Ro DK (2011) Lettuce costunolide synthase (CYP71BL2) and its homolog (CYP71BL1) from sunflower catalyze distinct regio- and stereoselective hydroxylations in sesquiterpene lactone metabolism. J Biol Chem 286:21601–21611
Jennewein S, Rithner CD, Williams RM, Croteau RB (2001) Taxol biosynthesis: taxane 13α-hydroxylase is a cytochrome P450-dependent monooxygenase. Proc Natl Acad Sci USA 98:13595–13600
Jennewein S, Rithner CD, Williams RM, Croteau RB (2003) Taxoid metabolism: taxoid 14β-hydroxylase is a cytochrome P450-dependent monooxygenase. Arch Biochem Biophys 413:262–270
Jensen K, Møller BL (2010) Plant NADPH-cytochrome P450 oxidoreductases. Phytochemistry 71:132–141
Keeling CI, Henderson H, Li M, Dullat HK, Ohnishi T, Bohlmann J (2013) CYP345E2, an antenna-specific cytochrome P450 from the mountain pine beetle, dendroctonus ponderosae hopkins, catalyses the oxidation of pine host monoterpene volatiles. Insect Biochem Mol Biol 43:1142–1151
Kim J, DellaPenna D (2006) Defining the primary route for lutein synthesis in plants: the role of Arabidopsis carotenoid β-ring hydroxylase CYP97A3. Proc Natl Acad Sci USA 103:3474–3479
Kim J, Smith JJ, Tian L, Dellapenna D (2009) The evolution and function of carotenoid hydroxylases in Arabidopsis. Plant Cell Physiol 50:463–479
Kim JE, Cheng KM, Craft NE, Hamberger B, Douglas CJ (2010) Over-expression of Arabidopsis thaliana carotenoid hydroxylases individually and in combination with a beta-carotene ketolase provides insight into in vivo functions. Phytochemistry 71:168–178
King AJ, Brown GD, Gilday AD, Larson TR, Graham IA (2014) Production of bioactive diterpenoids in the euphorbiaceae depends on evolutionarily conserved gene clusters. Plant Cell 26:3286–3298
Le-Huu P, Heidt T, Claasen B, Laschat S, Urlacher VB (2015) Chemo-, regio-, and stereoselective oxidation of the monocyclic diterpenoid β-cembrenediol by P450 BM3. ACS Catal 5:1772–1780
Le-Huu P, Petrović D, Strodel B, Urlacher VB (2016) One-pot, two-step hydroxylation of the macrocyclic diterpenoid β-cembrenediol catalyzed by P450 BM3 mutants. ChemCatChem 8:3755–3761
Li D, Ma Y, Zhou Y, Gou J, Zhong Y, Zhao L, Han L, Ovchinnikov S, Ma L, Huang S, Greisen P, Shang Y (2019) A structural and data-driven approach to engineering a plant cytochrome P450 enzyme. Sci China Life Sci 62:873–882
Linden H (1999) Carotenoid hydroxylase from Haematococcus pluvialis: cDNA sequence, regulation and functional complementation. BBA-Gene Struct Expr 1446:203–212
Maoka T (2020) Carotenoids as natural functional pigments. J Nat Med 74:1–16
Mau CJ, Karp F, Ito M, Honda G, Croteau RB (2010) A candidate cDNA clone for (−)-limonene-7-hydroxylase from Perilla frutescens. Phytochemistry 71:373–379
Misra RC, Sharma S, Sandeep Garg A, Chanotiya CS, Ghosh S (2017) Two CYP716A subfamily cytochrome P450 monooxygenases of sweet basil play similar but nonredundant roles in ursane- and oleanane-type pentacyclic triterpene biosynthesis. New Phytol 214:706–720
Moses T, Pollier J, Almagro L, Buyst D, Van Montagu M, Pedreno MA, Martins JC, Thevelein JM, Goossens A (2014) Combinatorial biosynthesis of sapogenins and saponins in Saccharomyces cerevisiae using a C-16alpha hydroxylase from Bupleurum falcatum. Proc Natl Acad Sci USA 111:1634–1639
Nadeau JA, Petereit J, Tillett RL, Jung K, Fotoohi M, MacLean M, Young S, Schlauch K, Blomquist GJ, Tittiger C (2017) Comparative transcriptomics of mountain pine beetle pheromone-biosynthetic tissues and functional analysis of CYP6DE3. BMC Genom 18:311
Nomura T, Magome H, Hanada A, Takeda-Kamiya N, Mander LN, Kamiya Y, Yamaguchi S (2013) Functional analysis of Arabidopsis CYP714A1 and CYP714A2 reveals that they are distinct gibberellin modification enzymes. Plant Cell Physiol 54:1837–1851
Paddon CJ, Keasling JD (2014) Semi-synthetic artemisinin: a model for the use of synthetic biology in pharmaceutical development. Nat Rev Microbiol 12:355–367
Park SY, Eun H, Lee MH, Lee SY (2022) Metabolic engineering of Escherichia coli with electron channelling for the production of natural products. Nat Catal 5:726–737
Pompon D, Louerat B, Bronine A, Urban P (1996) Yeast expression of animal and plant P450s in optimized redox environments. Methods Enzymol 272:51–64
Qi X, Bakht S, Qin B, Leggett M, Hemmings A, Mellon F, Eagles J, Werck-Reichhart D, Schaller H, Lesot A, Melton R, Osbourn A (2006) A different function for a member of an ancient and highly conserved cytochrome P450 family: from essential sterols to plant defense. Proc Natl Acad Sci USA 103:18848–18853
Qi DD, Jin J, Liu D, Jia B, Yuan YJ (2020) In vitro and in vivo recombination of heterologous modules for improving biosynthesis of astaxanthin in yeast. Microb Cell Fact 19(1):103
Quaderer R, Omura S, Ikeda H, Cane DE, Cane DE (2006) Pentalenolactone biosynthesis. Molecular cloning and assignment of biochemical function to PtlI, a cytochrome P450 of Streptomyces avermitilis. J Am Chem Soc 128:13036–13037
Quinlan RF, Jaradat TT, Wurtzel ET (2007) Escherichia coli as a platform for functional expression of plant P450 carotene hydroxylases. Arch Biochem Biophys 458:146–157
Ramirez AM, Saillard N, Yang T, Franssen MC, Bouwmeester HJ, Jongsma MA (2013) Biosynthesis of sesquiterpene lactones in pyrethrum (Tanacetum cinerariifolium). PLoS ONE 8:e65030
Renata H (2021) Synthetic utility of oxygenases in site-selective terpenoid functionalization. J Ind Microbiol Biotechnol. https://doi.org/10.1093/jimb/kuab002
Rouck JE, Biggs BW, Kambalyal A, Arnold WR, De Mey M, Ajikumar PK, Das A (2017) Heterologous expression and characterization of plant taxadiene-5alpha-hydroxylase (CYP725A4) in Escherichia coli. Protein Expr Purif 132:60–67
Schalk M, Croteau RA (2000) Single amino acid substitution (F363I) converts the regiochemistry of the spearmint (-)-limonene hydroxylase from a C6- to a C3-hydroxylase. Proc Natl Acad Sci U S A 97:11948–11953
Scheler U, Brandt W, Porzel A, Rothe K, Manzano D, Bozic D, Papaefthimiou D, Balcke GU, Henning A, Lohse S, Marillonnet S, Kanellis AK, Ferrer A, Tissier A (2016) Elucidation of the biosynthesis of carnosic acid and its reconstitution in yeast. Nat Commun 7:12942
Schifrin A, Litzenburger M, Ringle M, Ly TT, Bernhardt R (2015) New sesquiterpene oxidations with CYP260A1 and CYP264B1 from Sorangium cellulosum So ce56. ChemBioChem 16:2624–2632
Schoendorf A, Rithner CD, Williams RM, Croteau RB (2001) Molecular cloning of a cytochrome P450 taxane 10β-hydroxylase cDNA from taxus and functional expression in yeast. Proc Natl Acad Sci USA 98:1501–1506
Schrepfer P, Ugur I, Klumpe S, Loll B, Kaila VRI, Bruck T (2020) Exploring the catalytic cascade of cembranoid biosynthesis by combination of genetic engineering and molecular simulations. Comput Struct Biotechnol J 18:1819–1829
Seki H, Ohyama K, Sawai S, Mizutani M, Ohnishi T, Sudo H, Akashi T, Aoki T, Saito K, Muranaka T (2008) Licorice β-amyrin 11-oxidase, a cytochrome P450 with a key role in the biosynthesis of the triterpene sweetener glycyrrhizin. Proc Natl Acad Sci USA 105:14204–14209
Seki H, Sawai S, Ohyama K, Mizutani M, Ohnishi T, Sudo H, Fukushima EO, Akashi T, Aoki T, Saito K, Muranaka T (2011) Triterpene functional genomics in licorice for identification of CYP72A154 involved in the biosynthesis of glycyrrhizin. Plant Cell 23:4112–4123
Shang Y, Ma Y, Zhou Y, Zhang H, Duan L, Chen H, Zeng J, Zhou Q, Wang S, Gu W, Liu M, Ren J, Gu X, Zhang S, Wang Y, Yasukawa K, Bouwmeester HJ, Qi X, Zhang Z, Lucas WJ, Huang S (2014) Biosynthesis, regulation, and domestication of bitterness in cucumber. Science 346:1084–1088
Shibuya M, Hoshino M, Katsube Y, Hayashi H, Kushiro T, Ebizuka Y (2006) Identification of beta-amyrin and sophoradiol 24-hydroxylase by expressed sequence tag mining and functional expression assay. FEBS J 273:948–959
Sintupachee S, Promden W, Ngamrojanavanich N, Sitthithaworn W, De-Eknamkul W (2015) Functional expression of a putative geraniol 8-hydroxylase by reconstitution of bacterially expressed plant CYP76F45 and NADPH-cytochrome P450 reductase CPR I from Croton stellatopilosus Ohba. Phytochemistry 118:204–215
Song M, Kim AC, Gorzalski AJ, MacLean M, Young S, Ginzel MD, Blomquist GJ, Tittiger C (2013) Functional characterization of myrcene hydroxylases from two geographically distinct Ips pini populations. Insect Biochem Mol Biol 43:336–343
Sun W, Xue H, Liu H, Lv B, Yu Y, Wang Y, Huang M, Li C (2020) Controlling chemo- and regioselectivity of a plant P450 in yeast cell toward rare licorice triterpenoid biosynthesis. ACS Catal 10:4253–4260
Takase H, Sasaki K, Shinmori H, Shinohara A, Mochizuki C, Kobayashi H, Ikoma G, Saito H, Matsuo H, Suzuki S, Takata R (2016) Cytochrome P450 CYP71BE5 in grapevine (Vitis vinifera) catalyzes the formation of the spicy aroma compound (−)-rotundone. J Exp Bot 67:787–798
Tamura K, Seki H, Suzuki H, Kojoma M, Saito K, Muranaka T (2017) CYP716A179 functions as a triterpene C-28 oxidase in tissue-cultured stolons of Glycyrrhiza uralensis. Plant Cell Rep 36:437–445
Tian L, DellaPenna D (2004) Progress in understanding the origin and functions of carotenoid hydroxylases in plants. Arch Biochem Biophys 430:22–29
Tzin V, Snyder JH, Yang DS, Huhman DV, Watson BS, Allen SN, Tang Y, Miettinen K, Arendt P, Pollier J, Goossens A, Sumner LW (2019) Integrated metabolomics identifies CYP72A67 and CYP72A68 oxidases in the biosynthesis of Medicago truncatula oleanate sapogenins. Metabolomics 15(6):85
van Beilen JB, Holtackers R, Luscher D, Bauer U, Witholt B, Duetz WA (2005) Biocatalytic production of perillyl alcohol from limonene by using a novel Mycobacterium sp. cytochrome P450 alkane hydroxylase expressed in Pseudomonas putida. Appl Environ Microbiol 71:1737–1744
Walls LE, Malci K, Nowrouzi B, Li RA, d’Espaux L, Wong J, Dennis JA, Semiao AJC, Wallace S, Martinez JL, Keasling JD, Rios-Solis L (2021) Optimizing the biosynthesis of oxygenated and acetylated taxol precursors in Saccharomyces cerevisiae using advanced bioprocessing strategies. Biotechnol Bioeng 118:279–293
Walter RM, Zemella A, Schramm M, Kiebist J, Kubick S (2022) Vesicle-based cell-free synthesis of short and long unspecific peroxygenases. Front Bioeng Biotechnol 10:964396
Wang E, Wagner GJ (2003) Elucidation of the functions of genes central to diterpene metabolism in tobacco trichomes using posttranscriptional gene silencing. Planta 216:686–691
Wang H, Han J, Kanagarajan S, Lundgren A, Brodelius PE (2013) Trichome-specific expression of the amorpha-4,11-diene 12-hydroxylase (cyp71av1) gene, encoding a key enzyme of artemisinin biosynthesis in Artemisia annua, as reported by a promoter-GUS fusion. Plant Mol Biol 81:119–138
Yadav VG (2014) Unraveling the multispecificity and catalytic promiscuity of taxadiene monooxygenase. J Mol Catal B-Enzym 110:154–164
Yang LE, Huang XQ, Hang Y, Deng YY, Lu QQ, Lu S (2014) The P450-type carotene hydroxylase PuCHY1 from Porphyra suggests the evolution of carotenoid metabolism in red algae. J Integr Plant Biol 56:902–915
Yasumoto S, Fukushima EO, Seki H, Muranaka T (2016) Novel triterpene oxidizing activity of Arabidopsis thaliana CYP716A subfamily enzymes. FEBS Lett 590:533–540
Yasumoto S, Seki H, Shimizu Y, Fukushima EO, Muranaka T (2017) Functional characterization of CYP716 family P450 enzymes in triterpenoid biosynthesis in tomato. Front Plant Sci 8:21
Zhang N, Han ZT, Sun GL, Hoffman A, Wilson IW, Yang YF, Gao Q, Wu JQ, Xie D, Dai JG, Qiu DY (2014) Molecular cloning and characterization of a cytochrome P450 taxoid 9á-hydroxylase in Ginkgo biloba cells. Biochem Biophys Res Commun 443:938–943
Zhang J, Dai L, Yang J, Liu C, Men Y, Zeng Y, Cai Y, Zhu Y, Sun Y (2016) Oxidation of cucurbitadienol catalyzed by CYP87D18 in the biosynthesis of mogrosides from Siraitia grosvenorii. Plant Cell Physiol 57:1000–1007
Zhang J, Hansen LG, Gudich O, Viehrig K, Lassen LMM, Schrubbers L, Adhikari KB, Rubaszka P, Carrasquer-Alvarez E, Chen L, D’Ambrosio V, Lehka B, Haidar AK, Nallapareddy S, Giannakou K, Laloux M, Arsovska D, Jorgensen MAK, Chan LJG, Kristensen M, Christensen HB, Sudarsan S, Stander EA, Baidoo E, Petzold CJ, Wulff T, O’Connor SE, Courdavault V, Jensen MK, Keasling JD (2022) A microbial supply chain for production of the anti-cancer drug vinblastine. Nature 609:341–347
Zhao B, Lin X, Lei L, Lamb DC, Kelly SL, Waterman MR, Cane DE (2008) Biosynthesis of the sesquiterpene antibiotic albaflavenone in Streptomyces coelicolor A3(2). J Biol Chem 283:8183–8189
Zhao B, Lei L, Vassylyev DG, Lin X, Cane DE, Kelly SL, Yuan H, Lamb DC, Waterman MR (2009) Crystal structure of albaflavenone monooxygenase containing a moonlighting terpene synthase active site. J Biol Chem 284:36711–36719
Zhao S, Tuan PA, Kim JK, Park WT, Kim YB, Arasu MV, Al-Dhabi NA, Yang J, Li CH, Park SU (2014) Molecular characterization of carotenoid biosynthetic genes and carotenoid accumulation in Lycium chinense. Molecules 19:11250–11262
Zhou Y, Ma Y, Zeng J, Duan L, Xue X, Wang H, Lin T, Liu Z, Zeng K, Zhong Y, Zhang S, Hu Q, Liu M, Zhang H, Reed J, Moses T, Liu X, Huang P, Qing Z, Liu X, Tu P, Kuang H, Zhang Z, Osbourn A, Ro DK, Shang Y, Huang S (2016) Convergence and divergence of bitterness biosynthesis and regulation in cucurbitaceae. Nat Plants 2:16183
Zhu M, Wang C, Sun W, Zhou A, Wang Y, Zhang G, Zhou X, Huo Y, Li C (2018) Boosting 11-oxo-β-amyrin and glycyrrhetinic acid synthesis in Saccharomyces cerevisiae via pairing novel oxidation and reduction system from legume plants. Metab Eng 45:43–50
Zi J, Peters RJ (2013) Characterization of CYP76AH4 clarifies phenolic diterpenoid biosynthesis in the lamiaceae. Org Biomol Chem 11:7650–7652
Zwick CR, Renata H (2020) Harnessing the biocatalytic potential of iron- and alpha-ketoglutarate-dependent dioxygenases in natural product total synthesis. Nat Prod Rep 37:1065–1079
Acknowledgements
The authors thank the research and development platform provided by State Key Laboratory of Bioreactor Engineering, Shanghai Collaborative Innovation Centre for Biomanufacturing, Frontiers Science Center for Materiobiology and Dynamic Chemistry, East China University of Science and Technology, Shanghai.
Funding
This work was financially supported by the National Natural Science Foundation of China (Nos. 21922804 and 32271540), the National Key Research and Development Program of China (Grants 2019YFA0905000 and 2021YFC2102900), Program of Shanghai Academic Research Leader (21XD1400800), and the Fundamental Research Funds for the Central Universities (Grant No. 22221818014).
Author information
Authors and Affiliations
Contributions
ZZ drafted the manuscript; QW, YG, ZH, RH, AL, JX, and HY revised the manuscript. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
The authors approved the consent for publishing the manuscript.
Competing interests
The authors have no conflicts of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, Z., Wu, QY., Ge, Y. et al. Hydroxylases involved in terpenoid biosynthesis: a review. Bioresour. Bioprocess. 10, 39 (2023). https://doi.org/10.1186/s40643-023-00656-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40643-023-00656-1